The world’s gliding mammals are an extraordinary group of animals that have the ability to glide from tree to tree with seemingly effortless grace. There are more than 60 species of gliding mammals including the flying squirrels from Europe and North America, the scaly-tailed flying squirrels from central Africa and the gliding possums of Australia and New Guinea.
The oldest fossils proposed to be flying squirrels — the extinct genus Oligopetes — appear in the Oligocene and have been discovered in Europe and the Mediterranean. A further 13 extinct genera have been discovered from Miocene and Pliocene deposits. There appears to have been somewhat of a shift in the core of flying squirrels from Europe in the Oligocene and Miocene to Asia during the Miocene and more recent epochs. This shift in turn is reflected where we find the majority of flying squirrels today in southern Eurasia and South-East Asia.
So what are the origins and relationships which have helped to shape the current distributions of the species we see today? The arrow-tailed flying squirrels (Hylopetes) and the dwarf flying squirrels (Petinomys) with nine and eight species respectively, appear to be closely related and diverged from each other in the Pliocene approximately 2.2 million years ago.
It is thought that the North American flying squirrels of the genus Glaucomys diverged from the arrow-tailed flying squirrels and the dwarf flying squirrels some 29 million years ago, and at the same time traversed the once present land bridge between Siberia and Alaska. The ancestral stock of the North American flying squirrels probably migrated throughout North America during the
Apart from the ancestors of the North American flying squirrels there is one species of extinct Sciurion recorded from Miocene deposits in southwestern Saskatchewan in Canada. There are five species of the extinct genus Petauristodon from the late Miocene and early Pliocene from southern California and Florida. In addition there is one species of the extinct genus Miopetaurista from the Pliocene recorded from Florida. The fossil records of the North American flying squirrels that we see today are known only from Pleistocene and Holocene deposits where specimens have been obtained from localities including Bedford County, Pennsylvania; Eagle Cave, West Virginia; Augusta County, Virginia; and Robinson Cave and Baker Bluff, Tennessee.
Sometime in the late Miocene the Bering Strait opened to separate North America and Asia, thereby stopping further movement of gliding mammals. Movements of flying squirrels are likely to have required not only land but also a forested corridor. The south-west of North America appears to have been occupied by a dry woodland, chaparral, grassland and thorn forest, which seems to have been unsuitable for the Southern Flying Squirrel (Glaucomys volans). Once in North America the flying squirrels of the genus Glaucomys underwent a relatively rapid diversification in the early-to-middle Pleistocene. Two distinct lineages were identified within the Northern Flying Squirrel (G. sabrinus), a western lineage consisting of populations from western California, Oregon, and Washington, and a much larger eastern lineage comprised of populations from the remainder of the species range (North Carolina, West Virginia, Michigan, Utah, Alaska, eastern Washington, British Columbia and Alberta).
The giant flying squirrels of the genus Petaurista are most closely related to the North Chinese Flying Squirrel (genus Aeretes), the woolly flying squirrels (genus Eupetaurus) and the Eurasian flying squirrels from the genus Pteromys. It is thought that the giant flying squirrels diverged from the Eurasian flying squirrels about

They may also have expanded northward during the warmer periods of the middle Pleistocene. At the same time, the ancestor of the Eurasian flying squirrels may have extended their distribution to northern and central parts of Eurasia. The present distributions of Petaurista and Pteromys squirrels do not overlap except in parts of China and Japan. However, Pleistocene fossils of Petaurista squirrels have been recorded in Europe as well as China and Japan.
So far, Pleistocene fossils of the Siberian Flying Squirrel have been found only from Japan. However, it appears that prior to the divergence within Eurasian populations, the Hokkaido population of Siberian Flying Squirrels could have already been separated from the Eurasian populations, as there were land bridges between Hokkaido and Sakhalin islands and the Eurasian continent during most glaciations. Today the Siberian Flying Squirrel has the widest distribution of any flying squirrel, extending all the way from Japan in the east, through China and Russia and to Finland in the west.
The broad distribution of the giant flying squirrels has been profoundly influenced by the changing geology and associated climate through time. The Miocene was a period of warmer global climates than compared with the preceding Oligocene or the following Pliocene. These changes in climate were initiated in the Eocene about 50 mya when the then island continent of India crashed into Asia and resulted in an uplifting in the region which ultimately produced the Himalayas. The push of India into Eurasia has continued ever since and included a period of rapid uplifting in the Oligocene some 20 million years ago that resulted in the establishment of the Qinghai—Tibet Plateau, known as the ‘roof of the world’, some eight million years ago. These changes caused global cooling and resulted in subtropical regions retreating and triggered the development of large ice caps, and successive glacial and interglacial periods that contributed to the formation of natural geographic barriers. It seems that these changes forced the giant flying squirrel lineages in the region to migrate to their present geographic regions by the shifting of their forest habitat, and led to southwestern China and possibly northern Pakistan, becoming refuges for the woolly flying squirrels (Eupetaurus) and other specialised mammals that occur on each side of the Himalayas.
The ancestor of the eastern and western trans-Himalayan woolly flying squirrels diverged about 10.8 mya and migrated independently to their present localities near the end of the Miocene. During the glacial and interglacial periods of the Pliocene, the woolly flying squirrels adapted to the cold habitat of mountain valleys during the glaciations and further uplift of the Himalayas and Qinghai—Tibet plateau.

During the glacial periods of the Pliocene and Pleistocene the lower sea levels allowed land bridges to form that enabled the migration of animals and humans between the continents and islands. It was during these periods that land bridges were established between Korea and southern Japan and the northern end of Ryukyu Archipelago. The glacial periods also allowed the predecessors of the Taiwan Giant Flying Squirrel to migrate from southern China to Taiwan. Hainan Island, which lies off the south-east coast of China, joined with the mainland approximately 1.8 mya.
The ancestors of the Japanese Giant Flying Squirrel, Japanese Flying Squirrel and the Siberian Flying Squirrel most likely migrated through Korea or possibly China into southern Japan. There is also the possibility that the ancestors of the Japanese Flying Squirrel and the Siberian Flying Squirrel arrived into northern Japan through Sakhalin via eastern Russia. This journey probably occurred through land bridges formed around in the early to middle Pleistocene as a result of the lower sea levels. Fossil records of Petaurista before the Holocene period are very rare on the Japanese islands. The middle Pleistocene fossils of the Japanese Flying Squirrel have been found in two localities that are very close to each other, while the late Pleistocene fossils have been found in six localities suggesting this was probably the timing of the migration to Japan.
The present broad geographical distributions of the Spotted Giant Flying Squirrel, Red Giant Flying Squirrel and the Indian Giant Flying Squirrel appear to be the result of the tectonic events of the Pliocene—Pleistocene period and advancing and retreating ice as the climate fluctuated between cold and warm periods. The glacial periods of the Pliocene—Pleistocene resulted in reductions in sea level of more than 120 metres compared with the present sea level, giving flying squirrels access to the islands of the Sunda Shelf including Borneo, Sumatra, Java and numerous smaller islands from mainland South-East Asia through the Malay Peninsula, Thailand, Cambodia and Vietnam.
In addition to climate influences the oceanic tectonic movements during the Pliocene—Pleistocene period appear to have allowed the dispersal of the Red Giant Flying Squirrel and Spotted Giant Flying Squirrel across to the islands of the Sunda Shelf. Migration patterns suggest the dispersal hypothesis seems more appropriate to explain their current geographical distributions.
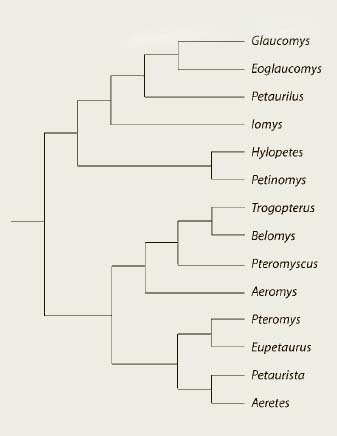
As we have seen, many different events have shaped the distribution and diversity of flying squirrels. But how are the different groups related to each other? Several attempts have been undertaken to assess the relationship of the existing genera of flying squirrels. A more recent analysis of all genera of flying squirrels, except the Namdapha Flying Squirrel (Biswamoyopterus biswasi), revealed there are three primary groups of flying squirrels. These include:
The exact position of Biswamoyopterus still remains to be resolved; however, it has a combination of characters that are present in other genera including Petaurista, Aeretes and Aeromys.

Thomas’s Flying Squirrel
Aeromys thomasi

Pakistan Woolly Flying Squirrel
Eupetaurus cinereus

Japanese Giant Flying Squirrel
Petaurista leucogenys

Complex-toothed Flying Squirrel
Trogopterus xanthipes