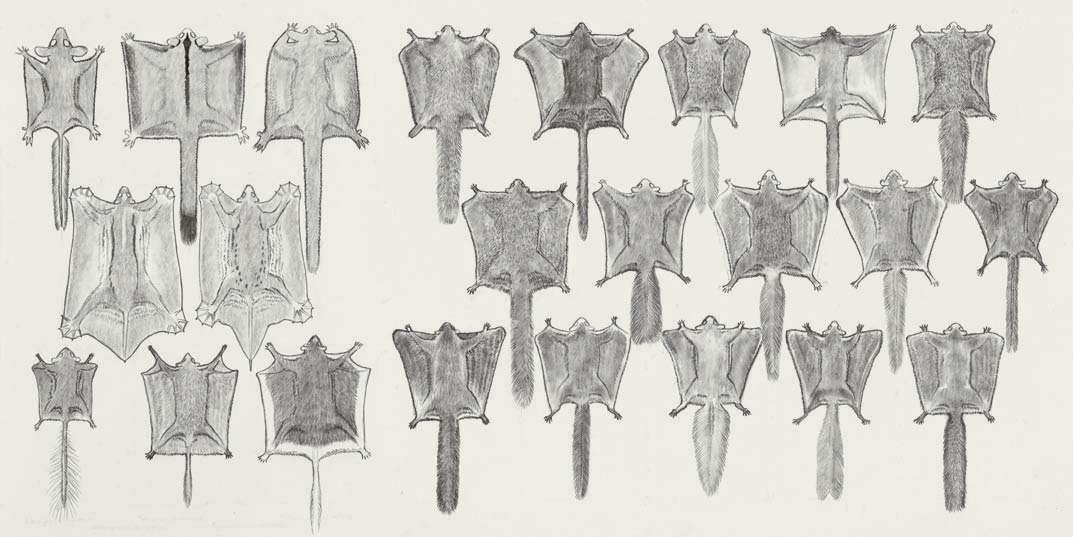
The world’s gliding mammals are an extraordinary group of animals that have the ability to glide from tree to tree with seemingly effortless grace. There are more than 60 species of gliding mammals including the flying squirrels from Europe and North America, the scaly-tailed flying squirrels from central Africa and the gliding possums of Australia and New Guinea.
The two things (which may be related) that link every species of gliding mammal together are their nocturnal behaviour and their ability to glide. Despite the diversity of their origins, the different groups of gliding mammals show a remarkable degree of convergence in relation to their gliding adaptations and behaviour. But how do these animals undertake this extraordinary method of locomotion and how far can they glide?
The patagia, or gliding membranes, of mammals consists of skin with two layers bound together tightly by connective tissue with muscles and nerves between. There are four types of patagia: the propatagium, the digipatagium (or dactylopatagium), the plagiopatagium and the uropatagium.
The propatagium (or neck membrane) attaches on each side of the neck and along the anterior edge of the forelimbs of the glider. This patagium is most developed in the colugos and little developed or absent in other gliding mammals. The digipatagia are found only in the two species of colugos and consist of membranes between each of the five digits on both the front and the hind feet. The plagiopatagium (or flank membrane) is the primary gliding membrane and is found in all species of gliding mammals. It extends between the forelimbs and hind limbs, and is under good muscular control. When the glider is climbing, resting or sleeping the plagiopatagium remains mostly hidden in the fur of the glider’s flanks. The exception to this can be found in the colugos, whose movements are somewhat inhibited by their overly developed flank membranes. The uropatagium (or tail membrane) extends between the posterior surface of the hind legs and the tail. Only the larger gliding mammals which weigh more than approximately 1 kilogram have these tail membranes. The uropatagium can range from completely enclosing the tail in the colugos, to including the proximal third of the tail in the large flying squirrels, to being absent or rudimentary in the smaller gliding mammals. Despite the variation in patagium design, its surface area in relation to body mass remains remarkably consistent with body size, regardless of the taxonomic group considered.

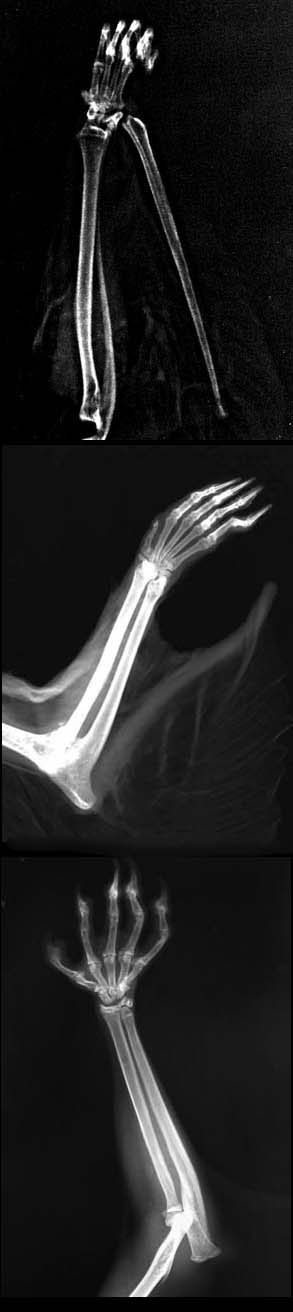
In the gliding possums of the genus Petaurus, the plagiopatagium extends from the joint of the second and third bones in its fifth digit of the front paw to the metatarsal region in the ankle of the foot. A similar arrangement is found in the plagiopatagium of the flying squirrels, although they possess a thin cartilaginous spur (called a styliform cartilage) that extends from the pisiform bone in the wrist. The extended cartilage increases the size of the patagium, stiffens and supports it and helps unfold its lateral leading edge. When not gliding, the cartilage is folded back and held against the forearm.
The scaly-tailed flying squirrels, which are unrelated to other flying squirrels, have an unciform cartilage that originates from the olecranon process of the ulna in the forearm (near the elbow) and helps support the leading edge of the plagiopatagium, effectively increasing the size of the patagium. The development of a styliform or unciform cartilage has allowed these species to evolve a wider membrane, independent of the length of the bones of the forelimbs.
The Greater Glider is unique among marsupial gliders because it has a very small accessory cartilaginous spur that extends from the olecranon process near the elbow. Its olecranon is also greatly elongated, extending considerably past its elbow joint. The animal glides by holding its paws under its chin so that its elbows extend out to the sides at right angles to the rest of its body, thus helping to increase the surface area of the plagiopatagium.
The curved styliform cartilage of the flying squirrels, unciform cartilage of the scaly-tailed flying squirrels, and inflected wrists of the petaurids and colugos form ‘winglets’ at the end of their forelimbs. There is remarkable similarity in the front appearance of the different groups of mammals as each group has independently come to the same evolutionary conclusion that winglets provide a valuable aeronautic advantage. Many birds, as well as modern aircraft, use these winglets to increase the efficiency and stability of their flight.

So how do these winglets work? They increase the effective width of the patagium and play an important role in the control and manoeuvrability of the glider. The near-vertical bending back of the winglet reduces turbulence at the leading edge of the patagium by redirecting the airflow from a downward directional force to a more lateral direction. This reduces the ‘drag’ that is created at the front edge of the gliding membrane where the high pressure below the wing ‘leaks out’ around the tip and produces a downward force to the wing, enabling the animal to more efficiently maintain lift. The winglets alter the airflow at the wing-tips and increase the effectiveness of the wing without materially increasing the wingspan. They smooth the airflow across the upper wing near the tip and reduce the strength of wing-tip vortices, thus improving the lift-to-drag ratio. The changes in airflow also enhance the stability of the glider by reducing ‘rolling’ and ‘yawing’ about the centre of gravity in the glider. Aeronautical engineers have determined by experiment that aircraft winglets can reduce drag by approximately 20 per cent and increase the lift-to-drag ratio by approximately 9 per cent.
There are other adaptations in the marsupial gliders of the genera Petaurus and Petauroides that improve their gliding ability. The skeletons of the smaller gliding possums show a slight elongation of the bones of the limbs and virtually no elongation of the tail, compared to similarly sized non-gliding possums. The Greater Glider too has developed comparatively longer bones in both its forelimbs and hind limbs, compared to its similarly sized non-gliding relatives, the ringtail possums.
Unlike the other marsupial gliders, the Feathertail Glider shows no marked elongation of its vertebrae or limb bones, suggesting that in such small animals the presence of the patagium alone is sufficient to provide the necessary increase in surface area, although this is no doubt assisted by its feather-like tail.
The forelimbs and hind limbs of the flying squirrels are also relatively longer than those of similarly sized non-gliding arboreal squirrels. As a result of the elongation of the bones in gliding mammals, the shape of the patagium has a roughly square shape compared with non-gliders which, if a patagium were stretched over their limbs, would have a rectangular shape. The square shape provides a proportionally greater surface area resulting in more lift and less drag than would a rectangular-shaped patagium. In addition, the square shape allows the glider to land at a relatively slower speed and with a high angle of attack, where the head is up and the feet are forward, instead of a low angle of attack, where the animal glides head first.
The shape of the tail of gliding mammals ranges from feather-like tails for the smallest species, to bushy, flattened tails or bushy, rounded tails with shortened fur for medium-sized mammals, to bushy, rounded tails with fluffy fur for the largest species.
Gliding mammals weighing approximately 30 grams or less, such as the marsupial Feathertail Glider (Acrobates), pygmy flying squirrels (Petaurillus) and pygmy scaly-tailed flying squirrels (Idiurus) typically have feather-like tails (known as ‘distichous tails’) which are dorso-ventrally flattened. The presence of a flattened tail appears to help the longitudinal (or ‘pitch’) control during gliding. The Feathertail Glider, for example, has a relatively narrow gliding membrane along the sides of its body, between the elbow and the knee. However, the effective patagium surface area is increased by long hairs that form a fringe along the margin of its tail.
The intermediate-sized gliders (weighing more than 30 grams to about 450 grams) have a range of tail types. The Arrow-tailed Flying Squirrel has a distichous tail, while the Hairy-footed Flying Squirrel has a tail that is more thickly furred but flattened. A number of species of the smaller marsupial gliders of the genus Petaurus and the Dwarf Scaly-tailed Flying Squirrel have a rounded, bushy tail.
Those species weighing more than about 450 grams, with the exception of the two species of colugos, have rounded, bushy tails, including the marsupial gliders (Petaurus and Petauroides), the giant flying squirrels (Petaurista), the large flying squirrels (Aeromys), and the woolly flying squirrels (Eupetaurus), and the larger scaly-tailed flying squirrels (Anomalurus).
Within those genera whose species have a large variation in body mass there is typically a degree of variation in tail morphology. For example, the smaller marsupial Sugar Glider has a less bushy tail than the larger heavier members of the genus. Similar observations have been made in the arrow-tailed flying squirrels (Hylopetes) and dwarf flying squirrels (Petinomys). The smallest dwarf flying squirrels, such as Temminck’s Flying Squirrel and Vordermann’s Flying Squirrel, have distichous tails, while heavier species, such as the Whiskered Flying Squirrel, have compactly furred but flattened tails. The still heavier Hagen’s Flying Squirrel and the Siberut Flying Squirrel, as well as the even larger Mindanao Flying Squirrel, have bushy, rounded tails.

Scaly-tailed flying squirrels have relatively long tails whose underside contains an area of rough, overlapping scales near the base, extending from one-fourth to one-third the length of the tail. It has been proposed that these scales are an ‘anti-skid’ device which the animal uses during landing or climbing. The potential for the scales to be actively involved in landing appears to be supported by observation of these species in the wild that, when the anomalures land, the tail makes a loud clacking sound as it slaps the tree trunk and digs into the bark. The tail appears to play only an incidental role in climbing as the animal does not put its whole body mass onto the scaly tail while climbing, but appears to use it when it is resting.

Feathertail Glider
Acrobates pygmaeus

Particolored Flying Squirrel
Hylopetes alboniger

Chinese Giant Flying Squirrel
Petaurista xanthotis

Pygmy Scaly-tailed Flying Squirrel
Idiurus zenkeri